Expresión génica, replicación y reparación del DNA en cromosomas multilaminares
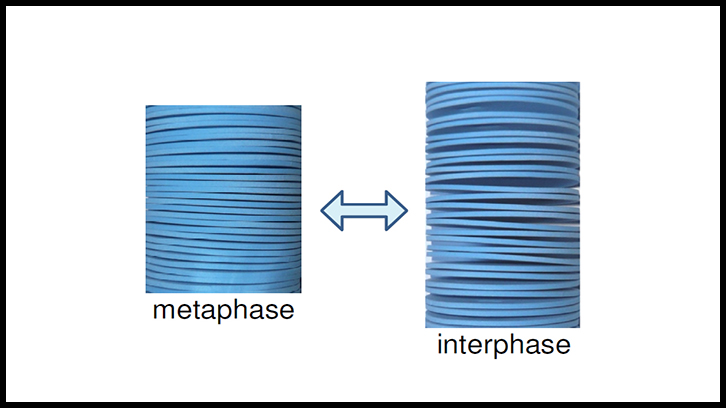
La condensación de los cromosomas y la división celular tienen lugar durante la mitosis, la última fase del ciclo celular. La transcripción se produce en todas las etapas (G1, S y G2) de la interfase, y la replicación del DNA ocurre solo durante la fase S. Conociendo estos tres procesos, los primeros estudios de microscopía electrónica mostraron la emanación de fibras de cromatina de cromosomas mitóticos y de núcleos interfásicos tratados con agua sin cationes. Desde entonces, la mayoría de los modelos cromosómicos propuestos tienen una relación directa con la geometría lineal de las fibras de cromatina. Sin embargo, en presencia de concentraciones de cationes correspondientes a la mitosis, los estudios de microscopía electrónica de transmisión, de microscopía de fuerza atómica y de crio-tomografía electrónica, realizados por investigadores del Laboratori de Cromatina del Departament de Bioquímica y Biologia Molecular de la UAB, mostraron que la cromatina emanada de los cromosomas mitóticos es una estructura plana que forma placas multilaminares compuestas de capas mononucleosomales.
Estos resultados llevaron a proponer el modelo de las placas delgadas, en el cual se considera que los cromosomas mitóticos condensados están formados por muchas capas de cromatina apiladas a lo largo del eje del cromosoma. Cada capa contiene aproximadamente 0.5 Mb de DNA en los cromosomas humanos. En tampones que contienen concentraciones de cationes correspondientes a la interfase, la cromatina emanada de los núcleos G1, S y G2 también tiene una morfología plana, pero las placas de los núcleos interfásicos tienen una baja tendencia a formar las estructuras multicapa observadas en los cromosomas mitóticos.
En un artículo de revisión e hipótesis publicado recientemente en FEBS Letters, se sugiere que durante la mitosis las capas de cromatina fuertemente apiladas inhiben la transcripción. Por el contrario, la accesibilidad a la cromatina aumenta enormemente cuando las capas no están tan densamente apiladas en la interfase, y el DNA puede interaccionar (por los dos lados de cada capa de cromatina) con las proteínas de la maquinaria de transcripción.
Los estudios de Captura de la Conformación de los Cromosomas a nivel de genoma completo (Hi-C) han demostrado que en la interfase hay muchos contactos entre las secuencias de DNA correspondientes a Dominios Asociados Topológicamente (TADs), que son considerados como subunidades funcionales de la cromatina. Dado que el tamaño de los TADs detectados en los análisis Hi-C de las células en interfase es similar a la cantidad de DNA en una capa de cromatina, se propone que cada capa corresponda a un TAD. Los bucles del filamento de cromatina propuestos a partir de los resultados obtenidos en los estudios de Hi-C pueden interpretarse como contactos producidos por el plegamiento del filamento dentro de las capas. En particular, los bucles enhancer-promotor requeridos para la expresión génica podrían ser preexistentes dentro de las capas. Durante la mitosis, los TADs aparentemente desaparecen porque las capas se apilan estrechamente y, en consecuencia, los contactos observados en los experimentos de Hi-C involucran secuencias más distantes en el genoma.
Según estudios anteriores, en primer lugar, es probable que la cromatina plana dentro de los cromosomas forme un helicoide continuo. La ausencia de entrecruzamientos topológicos entre las capas sucesivas de este helicoide puede facilitar la replicación del DNA. En segundo lugar, la cromatina plana tiene que ser localmente desestructurada para formar burbujas de replicación activas, y presumiblemente la replicación completa del cromosoma daría lugar a un doble helicoide sin entrecruzamientos topológicos. Esta estructura contiene dos copias de todas las secuencias de DNA originales en estrecha proximidad entre ellas, lo que puede facilitar la reparación de roturas de doble cadena por recombinación homóloga en cualquier región del genoma. Además, si se producen roturas de doble cadena, la red formada por la cromatina plana puede mantener los dos extremos del DNA en su lugar, evitando así las translocaciones cromosómicas, de una manera mejor que los filamentos de cromatina poco estructurados que normalmente se consideran en las publicaciones científica actuales.
Finalmente, la separación de los dos helicoides, por un mecanismo desconocido que presumiblemente requiere topoisomerasa II, producirá dos cromátidas hermanas compactas que pueden transferir de manera segura el DNA genómico a las células hijas.
Facultat de Biociències
Universitat Autònoma de Barcelona
Referencias
JR. Daban. Supramolecular multilayer organization of chromosomes: possible functional roles of planar chromatin in gene expression and DNA replication and repair. FEBS Letters 594 (2020) 395-411. https://febs.onlinelibrary.wiley.com/doi/10.1002/1873-3468.13724